Publié le 25 novembre 2016
Mis à jour le 18 mai 2026
Impacts du changement climatique : Eau et Biodiversité
Un indicateur est une information, associée à un phénomène, permettant d’en indiquer l’évolution dans le temps, de façon objective, et pouvant rendre compte des raisons de cette évolution. Dans cette fiche, les indicateurs s'intéressent à l'eau et la biodiversité.
Front d’expansion de la chenille processionnaire du pin
Cet indicateur rend compte de l’évolution, en France métropolitaine continentale, du front d’expansion de la chenille processionnaire du pin depuis les années 1970.
Responsable de l’indicateur : Jérôme Rousselet
Organisme propriétaire des données : INRAE (Institut National de Recherche pour l'agriculture, l'alimentation et l'environnement)
Données disponibles sur : Recherche Data Gouv entrepôt INRAE
Date de dernière actualisation de l’analyse technique et des données : 8 octobre 2024
La processionnaire du pin présente l'intérêt de produire un nid d'hiver qui est une structure très identifiable, visible de loin pendant plusieurs mois de l'année, et, pour un insecte, se prêtant très bien à la méthode dite du road sampling. La cartographie de sa distribution spatiale était assez grossière par le passé mais fournit néanmoins des données antérieures à l'expansion contemporaine servant aujourd'hui de référence (expansion observée en France et en Europe à partir de la fin des années 1980 / début des années 1990). Cette cartographie était obtenue par repérage visuel non exhaustif des nids par un opérateur expert mais sans méthode et effort d'échantillonnage clairement définis. L’observateur reportait ensuite manuellement ses observations sur une carte routière. En regroupant toutes ses observations sur plusieurs années, le CEMAGREF a ainsi produit une carte d'avancée maximale de l'insecte sur la période 1969-1979 (rapports internes, 1982, 2001).
Compte tenu de son intérêt pour l’étude du changement climatique, l’INRA a pris la décision en 2004 de constituer une base de données d'occurrence selon une méthode standardisée, reproductible par n'importe quel observateur, pour servir de référence à des comparaisons scientifiques sur le long terme. Il a également été fait le choix d'enregistrer des présences et des absences, ces dernières constituant le caractère unique des jeux de données produits. Pour se faire une grille de 8 x 8 km a été définie en se calant sur celle du réseau systématique du Département de la Santé des Forêts (DSF, Ministère en charge de l'Agriculture).
A partir des derniers signalements connus du DSF (uniquement des positifs enregistrés de façon opportuniste), une zone de prospection est définie à la marge de l'aire à chaque nouvelle campagne de cartographie. Les cellules sont prospectées en véhicule et considérées positives à partir du moment où au moins un nid y est observé, l'opérateur passant alors aux cellules voisines. Une fois l'ensemble des routes carrossables parcourues sans voir de nid (ou l'ensemble du territoire ayant pu être observé à l’œil nu ou aux jumelles), la cellule est déclarée négative à cet effort d'échantillonnage. Si une cellule du front anciennement positive est devenue négative, l'opérateur recherche une nouvelle cellule positive en amont du front. Toutes les cellules de 16x16 km dans l'aire de distribution de l'insecte en France ayant fait l'objet d'au moins un signalement entre 1981 et 2011, les cellules en arrière de la ligne de front définie selon la méthode énoncée ci-dessus sont considérées positives sans être reprospectées à chaque campagne.
Ce travail a été poursuivi par INRAE qui dispose maintenant de données collectées selon le même protocole s'étendant sur 20 ans au pas de temps de 5 ans. Le front d'expansion latitudinale a ainsi été enregistré pour les générations 2005-06, 2010-11, 2015-16 et 2020-21 (oeufs en année n et adultes en année n+1). La cartographie des fronts en montagne a pu être réalisée quant à elle sur une à deux générations selon les campagnes, et pour certaines régions ne pas être réalisées au même pas de temps ou au même grain. Le Massif central est ainsi disponible en 8x8 km en 2004-2006, 2014-2016, 2020-2021 et en 16x16 en 2011. Les Alpes et les Pyrénées le sont en 8x8 en 2015-16 et 2020-21 et en 16 x 16 en 2011.
Cet indicateur repose sur un lien de causalité démontré entre la réponse biologique (l'expansion) et la changement climatique au travers de la physiologie de l'insecte (températures requises pour son comportement alimentaire, et en conséquence de sa survie). Par contre, la vitesse d'expansion et la conquête vers le nord et en altitude des territoires devenus favorables à l'insecte avec le changement climatique est limitée par ses capacités de dispersion active (comportement et distance de vol des femelles), modulée par la structure du paysage (connectivité du milieu résultant de la distribution de ses arbres-hôtes et variant régionalement), et favorisée par des transports accidentels au-delà du front (dispersion passive). Par ailleurs, les réponses phénologiques de l'insecte au changement climatique sont susceptibles d'influencer ses réponses distributionnelles. Après chaque campagne de cartographie quinquennale en présence/absence, le DSF concentre son attention et son effort de signalements sur la zone de front défini par INRAE. Un nombre infime de cellules considérées négatives par INRAE ont finalement du être considérées positives suite à un signalment d'une autre source la même génération (en dehors de nouveaux foyers d'introduction découverts loin en aval du front).
L’utilisation d’indicateurs d’ordre biologique présente l’intérêt d’une approche intégrée des conséquences du changement climatique. Les contraintes thermiques sur le développement et la survie de l’insecte sont de plusieurs ordre, mais celle qui a constitué un facteur limitant sa distribution et rendre compte de ses possibilités d'expansion actuelle est celle qui conditionne les possibilités d'alimentation des larves entre octobre et mars (avec l’exigence d’une température du nid d’au minimum 9 °C durant le jour suivi d’une température de l’air supérieure à 0 °C la nuit suivante). Alors que ces contraintes forçaient par exemple en Région Centre l’insecte à stationner au sud de la Loire dans les années 1970, le réchauffement climatique les a progressivement levées permettant une expansion continue vers le Nord (augmentation des températures minimales d'octobre à mars d'environ 1°C entre les 15 années avant 1990 et les 15 années après). A l’inverse, en Tunisie, des signes de rétraction de sa limite sud sont observés avec l'augmentation des températures estivales. INRAE dispose d’un modèle prévisionnel permettant de simuler l’expansion selon différents scenarii climatiques pour le futur. Un argument supplémentaire réside dans l’impact sanitaire conséquent (urtication/allergies/choc anaphylactique) pour l’Homme et les animaux domestiques.
Etre réactualisé en 2025-2026. Explorer les possibilités d'une cartographie à l'échelle continentale en 16 x 16 km régulièrement actualisée en recourant à de l'intelligence artificielle pour analyser des images disponibles en ligne. Pour la France, on peut s'attendre à la colonisation de la majeure partie du territoire métropolitain à l'horizon d'une quinzaine d'années.
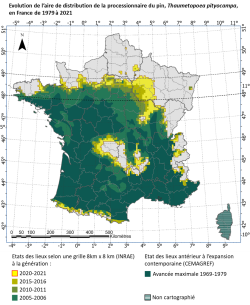
La carte de son aire de distribution en France, réactualisée tous les 5 ans depuis 2005, montre l’expansion latitudinale de la processionnaire du pin entre sa limite nord maximale sur la période 1969-1979 (données CEMAGREF) et le front de colonisation enregistré à l’hiver 2020-2021 (dernière mise à jour INRAE). Cet insecte originaire du bassin méditerranéen présente, contrairement à la plupart des insectes de zone tempérée, un développement larvaire hivernal. Il se trouve, pendant ce stade, favorisé par une augmentation de la température hivernale qui régule ses chances de survie. En l'occurrence, les chenilles ont besoin pour pouvoir se nourrir d'une température d'au moins 9°C dans leur nid durant le jour suivi d'une température de l'air supérieure à 0°C la nuit suivante (durant laquelle elles sortent s'alimenter). Alors que ces contraintes thermiques forçaient par exemple en Région Centre l’insecte à stationner au sud de la Loire dans les années 1970, le réchauffement climatique les a progressivement levées permettant une expansion continue vers le Nord. Les populations situées à l’Est et à l’Ouest du Massif central, séparées à l’origine, se sont également rejointes au nord de ce massif. Une progression similaire est visible en altitude (Alpes, Pyrénées, Massif central). La progression la plus rapide du front Nord a eu lieu dans la région Grand Est (en Champagne).
Températures minimales d'octobre à mars à Orléans avant et après le début de l'évolution du front de colonisation
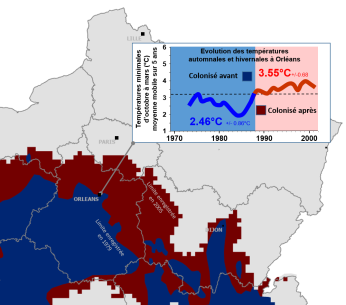
Orléans est une ville qui était située en limite des territoires colonisés et non colonisés tels que décrits en 1979. Une succession d'hivers chauds amenait la processionnaire du pin jusqu'à Orléans tandis qu'une succession d'hivers froids la cantonnait un peu plus au sud (expansions-rétractions avec une limite maximale au niveau de cette ville correspondant au point le plus au nord de la vallée de la Loire). Fin des années 1980/début des années 1990, une expansion incessante vers le nord a commencé à être observée. Dans le même temps, on assistait à une évolution des températures minimales d'octobre à mars qui sont corrélées aux températures qui déterminent pour les chenilles les possibilités de s'alimenter ou non, et donc leur survie.
Indice de sécheresse annuelle en métropole
Cet indicateur rend compte de la sécheresse agricole (ou édaphique), en France métropolitaine, calculé comme le pourcentage en moyenne sur l’année de la surface du territoire où l’on a observé des conditions anormales d’humidité du sol.
Responsable de l’indicateur : Simon Mittelberger
Organisme propriétaire des données : Météo-France
Date de dernière actualisation de l’analyse technique et des données : 15 octobre 2025
L’indice reproduit est un indicateur de la sécheresse agricole (ou édaphique) en France métropolitaine issu des travaux du projet Climsec (Soubeyroux et al, 2012), basé sur l’indice d’humidité du sol, Soil Wetness Index (SWI).
Il est calculé comme le pourcentage en moyenne sur l’année de la surface du territoire où l’on a observé des conditions anormales d’humidité du sol.
Le critère appliqué est le suivant : moyenne sur les 12 mois de l’année de la surface du territoire où l’humidité du sol est inférieure au seuil des 10 % des humidités du sol les plus faibles que l’on ait observées pour le même mois et au même lieu sur la période 1961-1990 (fréquence d’apparition de l’événement de sécheresse supérieure à 10 ans).
La réanalyse hydrométéorologique de la chaîne Safran-Isba-Modcou (SIM) sur la période 1958-2008 (Vidal et al., 2009) a permis dans le cadre du projet ClimSec (soutenu par la Fondation Maif) de disposer d’une base de données homogène à haute résolution spatio-temporelle sur la France pour caractériser les déficits en eau des différentes composantes du cycle de l’eau.
Des indicateurs standardisés, basés sur les l’indice d’humidité des sols (SSWI) ont été définis et utilisés pour décrire la typologie des sécheresses dans le climat actuel (Vidal et al., 2009). Les données de base sont issues de la chaîne SIM, opérationnelle à Météo-France depuis 2003. Les données sont disponibles depuis 1959.
Sous l’effet des changements climatiques, la fréquence et l’ampleur des sécheresses vont très certainement s’accroître. La France est particulièrement concernée par ce risque, comme ont pu le montrer les études sur les évolutions attendues du cycle hydrologique, mais l’étude des sécheresses nécessite de s’accorder sur des définitions communes des différents types de phénomène – météorologique, agricole et hydrologique – et de définir des indices normalisés pour les caractériser.
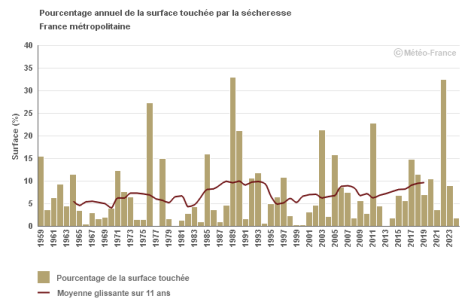
L’analyse du pourcentage annuel de la surface touchée par la sécheresse des sols depuis 1959 permet d’identifier les années ayant connu les événements les plus sévères comme 1976, 1989, 2003, 2011 et 2022
L’évolution de la moyenne décennale montre l’augmentation de la surface des sécheresses passant de valeurs de l’ordre de 5 % dans les années 1960 à plus de 10 % de nos jours.
Evolution de la date de migration de certains oiseaux
4,8 jours En moyenne, au cours de la dernière décennie, les oiseaux migrateurs transsahariens observés à la Pointe de Grave pour leur migration prénuptiale reviennent 4,8 jours plus tôt que lors des premières observations dans les années 1980-1990.
Cet indicateur présente l’avancée des dates d’arrivée printanières des migrateurs sur leurs zones de nidification, à partir d’effectifs nationaux ou régionaux. Le site retenu est la Pointe de Grave (33), à l’extrémité nord de la pointe du Médoc. Ce site concentre un flux important de migrateurs à la migration prénuptiale.
Responsable de l’indicateur : Jérémy DUPUY, LPO (Ligue pour la Protection des Oiseaux)
Organisme propriétaire des données : LPO
Traitement des données : DGEC
Date de dernière actualisation de l’analyse technique et des données : avril 2026
Le site de la Pointe de Grave (33), situé à l’extrémité nord de la pointe du Médoc, concentre un flux important de migrateurs à la migration prénuptiale. Dans le cadre de cet indicateur, les espèces dites migratrices transsahariennes ont été sélectionnées afin d’apporter une homogénéité dans les stratégies de migration déployées par les migrateurs. En effet, les migrateurs transsahariens sont en grande majorité des migrateurs totaux, c’est-à-dire que l’ensemble de la population nicheuse européenne migre au sud du Sahara pour hiverner. Les données utilisées pour calculer cet indicateur ont été récoltées lors du suivi de la migration active mené dans le Sud-Ouest de la France par la LPO Délégation territoriale Aquitaine : dans le département de la Gironde (33), à la Pointe de Grave pour la migration prénuptiale. Durant ce suivi, des observateurs professionnels assistés par des écovolontaires, observent, identifient et comptabilisent les oiseaux en migration active, du lever au coucher du soleil. Au printemps, le suivi se déroule du 1er mars au 31 mai (avec cependant des différences selon les années) et a débuté en 1986 de manière partielle. Les données récoltées se décrivent sous la forme de totaux journaliers pour chaque espèce observée en migration active. Ainsi, chaque année, une date moyenne de passage est obtenue pour les espèces qui nous intéressent. Pour cet indicateur, la date moyenne a été privilégiée, car elle apparait comme la plus robuste (Moussus et al. 2010).
Afin de s'affranchir des fortes variations interannuelles constatées, les courbes représentées sur le premier graphique font apparaître la moyenne décennale des dates moyennes annuelles de passage de chaque espèce. Le deuxième graphique représente la date moyenne annuelle de passage pour l'ensemble des 16 espèces observées.
Les espèces retenues sont celles pour lesquelles :
1/ la série d’observations est suffisamment longue pour être significative (jeu de données dépassant parfois 40 ans)
2/ le recensement, régional ou national, est de très bonne fiabilité
Dans l’étude du vivant, les oiseaux tiennent une place particulière. Sous nos latitudes et parce qu’ils sont diurnes pour la plupart, assez peu nombreux en termes de diversité (quelques centaines d’espèces au plus), facilement visibles (plumages colorés des mâles) et audibles (chants, cris), ils sont parmi les espèces les plus étudiées par les écologues, mais aussi parmi ceux qui touchent le plus le grand public. Les conséquences biologiques du changement climatique sur les oiseaux sont désormais bien connues (Møller et al. 2010). Une des réponses les plus documentées repose sur des décalages temporels dans la phénologie des espèces. Parmi ces décalages, l’avancée des dates d’arrivées printanières des migrateurs sur leurs zones de nidification est bien détaillée (Sokolov et al. 1998; Lehikoinen et al. 2004 ; Sparks et al. 2005 ; Jonzén et al. 2006 ; Zalakevicius et al. 2006).
Les données présentées lors de l’étude n’auront a priori pas à subir, à court terme, de modifications de la procédure de récolte, ni de leur traitement statistique.
Date de passage au printemps de 16 espèces de migrateurs transsahariens
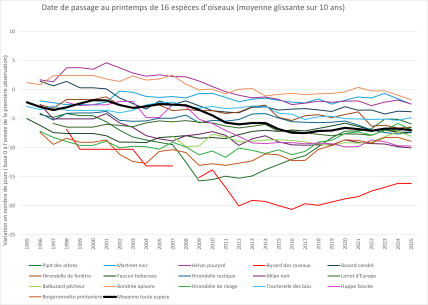
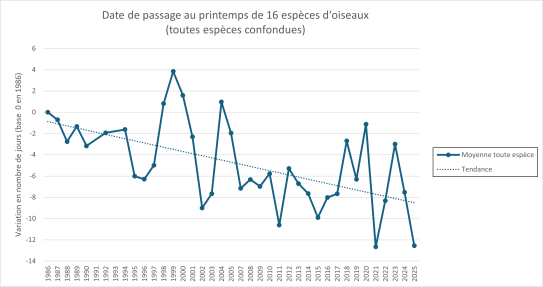
Les 16 espèces ne répondent pas de la même manière au réchauffement climatique. Ainsi, le décalage temporel atteint neuf jours en moyenne pour le busard des roseaux tandis qu'il est d'environ un jour pour le martinet noir, le héron pourpré et le loriot d'Europe. Les dates de migrations peuvent également fortement varier d'une année sur l'autre. Toutefois, toutes les espèces de migrateurs transsahariens observés présentent en moyenne une date retour plus précoce depuis les trois dernières décennies.
Températures moyennes des eaux du Léman
Cet indicateur rend compte de l’évolution de la température en moyenne annuelle des eaux du Léman à la surface et en profondeur.
Personne contact : Giuilia MARTI
Organisme propriétaire des données : Commission Internationale pour la Protection des Eaux du Léman – CIPEL – www.cipel.org
Date de dernière actualisation de l’analyse technique et des données : novembre 2024
La température moyenne par niveau est calculée à partir de la moyenne mensuelle, puis la moyenne des températures moyennes mensuelles. Lorsque les données ne sont pas disponibles, elles ont été implémentées à l'aide du modèle SIMSTRAT (Goudsmit et al., 2002).
Pour la période 1992 - 2023, les données sont issues de la station de référence SHL2 située au milieu du lac, dans la zone de profondeur maximale (46° 27’ 05.1"N 6° 34’ 23.8"E).
Les données recueillies permettent d’analyser les tendances à long terme de la température de la masse d’eau.
La température moyenne est calculée à partir de 18 à 21 relevés effectués chaque année pour chaque profondeur mesurée.
Le suivi de l’évolution de la température des eaux du Léman permet d’estimer l’ampleur du réchauffement du lac en lien avec le changement climatique, comme :
• le brassage hivernal des eaux, qui permet la réoxygénation des eaux du fond ;
• le développement du phytoplancton qui, s’il est trop important, peut causer des nuisances pour les usagers du lac (productions massives, baisse de la transparence, colmatage des filets de pêche, etc.) ;
• la reproduction de certaines espèces de poissons comme l’omble chevalier et le corégone dont la température optimale pour la reproduction doit être 8°C.
Évolution des températures de surface (0-10m) des eaux du Léman
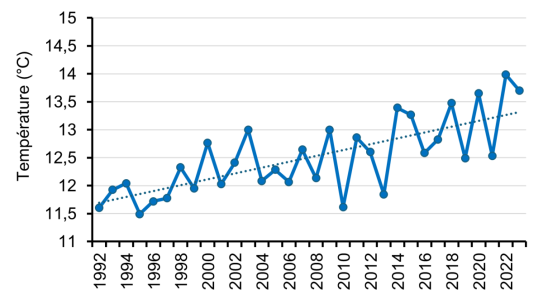
La température moyenne annuelle de surface (0-10 m) présente une nette tendance à l’augmentation (+1.3°C en moyenne sur la période 1992-2022, soit environ +0.05 °C/an).
Évolution de la température du fond du lac (300 - 309 m)
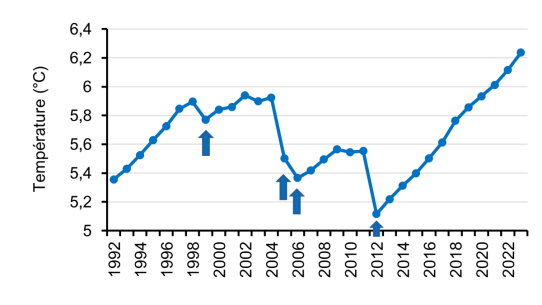
Au fond du lac, la température augmente continuellement en l’absence de brassage hivernal complet et chute brutalement lors des brassages hivernaux complets (1999, 2005, 2006 et 2012). Depuis le dernier brasage hivernal complet en 2012, la température au fond du lac a augmenté de +1.1 °C, soit 0.1 °C/an.
Le lac devient plus chaud sur une plus grande période de l’année. Cette augmentation peut impacter la reproduction de certaines espèces de poissons, ainsi que le développement du phytoplancton. L’algue filamenteuse Mougeotia gracillima, généralement observée en période automnale dans le Léman, peut dorénavant présenter des biomasses importantes en été.
Voir aussi
Impacts du changement climatique : Littoral et Milieu marin
Publié le 10 janvier 2017
Mis à jour le 21 avril 2026
Impacts du changement climatique : Agriculture et Forêt
Publié le 09 janvier 2017
Mis à jour le 24 février 2026